Lid geworden op: do jul 21, 2011 3:38 pm
Berichten: 9579
|
De eerste pagina van het gepubliceerde onderzoek: https://search.proquest.com/openview/6e ... &cbl=38594En nog een link: https://www.appliedanimalbehaviour.com/ ... 68-1591(18)30035-2/fulltext Ha, en hier vindt u de volledige tekst: https://onlinelibrary.wiley.com/doi/ful ... /zoo.21400Cognitive judgement bias is associated with frequency of anticipatory behavior in bottlenose dolphins Isabella L. K. Clegg, Fabienne Delfour
First published: 31 January 2018 |https://doi.org/10.1002/zoo.21400Abstract Many animals display a suite of increased vigilance and/or activity responses in relation to upcoming events, termed “anticipatory behavior.”
Anticipatory behavior toward positive events has been suggested as a cross‐species measure of affective state as it likely reflects the balance of the reward‐sensitivity system: various studies suggest that animals in poorer welfare situations show higher or excessive levels of anticipation for positive events.
Another tool for evaluating animals' affective state is cognitive bias testing, and although it has been attempted, a link has not yet been made between cognitive bias and anticipatory behavior levels.
Bottlenose dolphins (Tursiops truncatus) in captivity increase the performance of behaviors such as surface‐looking and spy‐hopping in anticipation of training sessions during which food is provided.
In this study we measured anticipatory behavior frequency in bottlenose dolphins prior to positive reinforcement training sessions, and assessed whether frequency of anticipatory behavior correlated with their performance on cognitive bias tasks.
We found that higher frequencies of anticipatory behavior for training sessions was significantly associated with more pessimistic judgements in cognitive bias tests, supporting previous findings linking higher reward sensitivity with negative affective states.
Anticipatory behavior is an easily measured activity and could represent a welfare indicator in dolphins as well as other animals in captive environments. 1 INTRODUCTIONAnticipatory behavior describes the activity performed by an individual in expectation of a predictable upcoming event (Spruijt, van den Bos, & Pijlman, 2001).
The ability to anticipate predictable events extends to many species, and in general anticipatory behavior is documented as increased activity, vigilance and/or increased transitions from one behavior to another (van den Bos, Meijer, van Renselaar, van der Harst, & Spruijt, 2003; van der Harst, Baars, & Spruijt, 2003).
Anticipatory activity has been most frequently described in relation to expected food events (Mistlberger, 2009; Storch & Weitz, 2009), but animals' anticipation has also been shown before access to play opportunities (Anderson, Yngvesson, Boissy, Uvnäs‐Moberg, & Lidfors, 2015), enriched housing (van der Harst, Fermont, Bilstra, & Spruijt, 2003), sexual interactions (van der Harst, Fermont, et al., 2003), and positive human‐animal interactions (Krebs, Torres, Chesney, Moon, & Watters, 2017).
Anticipatory behavior has recently garnered interest in animal welfare research (van der Harst & Spruijt, 2007; Watters, 2014) because it is thought to reflect underlying affective states (combinations of discrete emotions, Mendl et al., 2010), through its links with the reward sensitivity system (Spruijt et al., 2001; van der Harst & Spruijt, 2007).
A handful of studies have demonstrated that animals in poorer welfare conditions show a significantly higher level of anticipation to positive events than conspecifics in better welfare conditions (e.g., rats, Rattus norvegicus, in standard versus enriched cages, van der Harst, Baars, et al., 2003; van der Harst, Fermont, et al., 2003; or social isolation versus group housing, van den Berg et al., 1999).
Animals experiencing negative affective states place more “incentive value” on acquiring positive rewards, and thus anticipate their arrival more intensely (e.g., a hungrier animal will place more incentive value on a food item, Spruijt et al., 2001). However the link between anticipatory behavior and affective state may not be a straightforward linear relationship.
Farmed silver foxes (Vulpes vulpes) increased anticipatory as well as stereotypical behaviors before a positive reward, but did not show stereotypical behaviors before negative events (Moe, Bakken, Kittilsen, Kingsley‐Smith, & Spruijt, 2006), while rats in depressed‐like states did not anticipate positive rewards at all (von Frijtag et al., 2000).
More work on this topic is necessary to understand the underlying motivational systems, but a theoretical model of the intensity of anticipatory behavior suggests intensity will increase with poorer welfare up until the point of chronic stress, after which it drops dramatically (van der Harst & Spruijt, 2007; Watters, 2014).
Another recently investigated and promising welfare assessment tool is cognitive bias testing (Mendl, Burman, Parker, & Paul, 2009; Wichman, Keeling, & Forkman, 2012).
Cognitive biases show the effect of emotions on an individual's cognitive functioning, and tests in many species have shown that animals in poorer welfare conditions judge ambiguous cues more pessimistically, and vice versa (latest reviews by Baciadonna & McElligott, 2015; Roelofs, Boleij, Nordquist, & van der Staay, 2016). Scientists in this field are becoming more interested in the effects of spontaneous behavior on the results of bias testing (Wichman et al., 2012), demonstrated by recent evidence supporting a link between affiliative social behavior and cognitive biases.
A study on canaries (Serinus canaria) found pair‐housing led to more optimistic judgements (Lalot, Ung, Péron, D'Ettorre, & Bovet, 2017), and bottlenose dolphins conducting more synchronous swimming also judged more optimistically (Clegg, Rödel, & Delfour, 2017).
Although one study with laying hens (Gallus gallus) correlated anticipatory behavior frequency to cognitive bias and found no associations, they suggested some methodological problems and recommended further investigation (Wichman et al., 2012).
Cognitive bias testing is time‐intensive due to the training process but in general is an accurate test of animals' affective state (Mendl et al., 2009; Roelofs et al., 2016), and anticipatory behavior is simpler and quicker to measure but thus far needs more evidence supporting links to affective state (Watters, 2014).
Understanding the relationship between these two welfare tests would be beneficial, as it would allow scientists and animal managers who are currently limited by the tests' disadvantages to use the most appropriate, dependent on the settings.
To examine the potential links between anticipatory behavior frequency and cognitive bias, we measured the frequency of anticipatory behaviors of captive bottlenose dolphins toward upcoming training sessions and correlated them to the animals' results from cognitive bias tests.
Previous studies suggest dolphins view the multiple daily training sessions positively as the sessions provide food, cognitive stimulation, and anticipatory behaviors are exhibited prior to sessions (Brando, 2010; Clegg, Rödel, Cellier, et al., 2017; Jensen, Delfour, & Carter, 2013; Laule, Bloomsmith, & Schapiro, 2003; Miller, Mellen, Greer, & Kuczaj II, 2011).
Therefore we hypothesized that based on the reward‐sensitivity theory (Spruijt et al., 2001), dolphins showing the higher frequencies of anticipatory behavior would also make more pessimistic judgements in the cognitive bias task.
2 MATERIALS AND METHODSFor further information on methodology, equipment, and test protocols please refer to Clegg and co‐authors' study (Clegg, Rödel, & Delfour, 2017).
2.1 Study animals and facility
Eight Atlantic bottlenose dolphins (Tursiops truncatus) housed at Parc Astérix (Plailly, France) were used in this study (four males, four females, age range: 4–43 years).
The dolphins participated in multiple daily training sessions, which could involve tasks conditioned using positive reinforcement (Brando, 2010) such as medical training, show practice, novel behaviors, free‐feeds, and play sessions. During the experiment, training sessions began with the trainers approaching the outside beach area and occurred at regular intervals throughout the day.
Furthermore, acoustic and visual environmental cues (e.g., trainers preparing and setting up the food buckets) allowed the animals to predict the start time of the upcoming training session, as was shown in a study on anticipatory behavior of training sessions at Parc Asterix (Jensen et al., 2013), and similar to other zoo animals (Krebs et al., 2017; Watters, 2014).
Since this situation had occurred for the last several years, the dolphins had been conditioned to cues signalling the imminent start of training sessions.
Fixed feeding schedules and predictable environmental cues have been shown to elicit anticipatory behavior in many species in zoo and other captive settings (Krebs et al., 2017), and such classical conditioning, that is, pairing experimental cues to rewards, has also been deliberately applied in other anticipatory behavior studies (e.g., Wichman et al., 2012).
The park was closed to the public for the duration of the experiment.
2.2 Cognitive bias testing
2.2.1 Test protocol: Judgement bias task from Clegg, Rödel, and Delfour (2017)
Individual cognitive biases were measured using a spatial judgement Go/Go task adapted by Clegg et al. (2017).
A three‐step procedure was designed with conservative criteria that had to be fulfilled in order to pass to the next stage: first the animals were taught the task behavior, which was to touch a target on the other side of the pool and return to the trainer.
Once the animal touched the target with their rostrum, a whistle (“bridge”) was blown by the target‐holder which indicated to that the correct behavior had been performed and they should return to the trainer.
In Step 2, repeated trials were conducted where the target was placed on either of the extreme positions (far left or right of an arc, see Figure 1 in Clegg, Rödel, Delfour, et al., 2017), which were differentially rewarded with either a herring, applause and rubs (the “Positive” [P] cue reward) or only applause and eye contact (the “Less‐positive” [LP] cue reward).
The animal care staff at Parc Astérix confirmed that all dolphins would perceive the Positive and the Less‐positive rewards as representing larger and smaller reinforcement, respectively. Herring is larger and has a higher fat content than the other types of fish fed at Parc Astérix (Fisher, Stewart, Kastelein, & Campbell, 1992), and thus was expected to be a stronger primary reinforcer.
Although dolphins react positively to applause and eye contact, this is considered a secondary reinforcement compared to food rewards (Brando, 2010; Neto, Silveira, & dos Santos, 2016).
In order to complete Step 2 and pass to the final testing phase of Step 3, evidence was needed that the dolphins were discriminating between the extreme cues: this would be assessed from the recorded return times to the trainer, with the criteria that they must return ≥2 s faster from the Positive than the Less‐positive cue (Clegg, Rödel, Delfour, et al., 2017).
The animals were not separated from their regular social groups during testing (advantages discussed in Roelofs et al., 2016), in order to avoid inducing stress from social isolation, which has often been recorded with dolphins (e.g., Waples & Gales, 2002).
The test dolphin would undergo the test while the other dolphins would remain with their trainers in other parts of the pool (no gates used, see Figure 1 in Clegg, Rödel, Delfour, et al., 2017 for layout). Interference from the other dolphins, and thus the possibility of social learning, was limited as much as possible in the methods: a visual cue, the target, was chosen (as opposed to a sound, which all dolphins would hear); some dolphins were always taken to the inside pool, and the rest stayed on the opposite side of the pool to the test, and thus not in the visual field of the test dolphin.
All other animals were well‐reinforced with food for staying with their trainers and not leaving to go to the test area.
Lastly, if another dolphin did enter the test area during a trial, it would be stopped and repeated once the animal had left.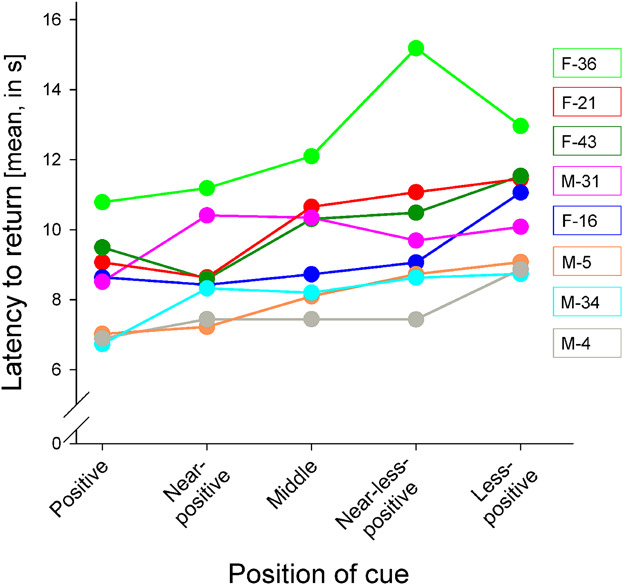 Figure 1Individual dolphins' absolute latencies (in s) to return from the conditioned (Positive and Less‐positive) and ambiguous (Near‐positive, Middle, Near‐less‐positive) cues. Figure 1Individual dolphins' absolute latencies (in s) to return from the conditioned (Positive and Less‐positive) and ambiguous (Near‐positive, Middle, Near‐less‐positive) cues.
Labels on the right hand side indicate the sex (M/F) and age of each of the eight dolphins.
When all animals' data were averaged, the latency to return from the Positive versus the Less‐positive cue was 2.0 s (min: 1.5 s, max: 2.4 s) (Clegg, Rödel, Delfour, et al., 2017), demonstrating that the dolphins had continued to differentiate between the two conditioned cues Once consistent differences in the speed to return to the trainer were seen (criteria: ≥2 s faster to return from the Positive than the Less‐positive cue), the animal progressed to Step 3, where the ambiguous cues were presented: the target was placed at three points along an arc shape between the P and LP positions.
The ambiguous cue positions were termed Near‐positive (NP), Middle (M), and Near‐less‐positive (NLP), and were equidistant from the start/finish position of the animal, and from each other. Within Step 3, each animal completed one cognitive bias test session per day (12 trials) for three consecutive days (total of 36 trials per animal), and participated in other daily training sessions as usual.
In each set of trials each ambiguous cue (NP, M, NLP) was presented once and the P and LP positions made up the other nine trials, following a pseudorandom order where a maximum of two positively (i.e., P) or less‐positively reinforced cue positions (i.e., NP, M, NLP, LP) could follow one another.
The reward given when the target was at the ambiguous positions was applause and eye contact, and the fact that the whistle was blown when the target was touched continued to let the animal know it had performed the correct behavior (Laule et al., 2003).
The return times to the trainer from each of the cues were recorded from the video footage.
The full tests were completed by each animal during January and February 2016, and the fact that all animals progressed to Step 3 (i.e., showed ≥2 s difference between the Positive and Less‐positive cue) validated our choice of reinforcement for these two extreme cues.
2.2.2 Cognitive bias response variables
The overall response variable reflecting cognitive bias was the dolphins' return latency (in s) from the target to the trainer, where it would receive the reward (the same as in Clegg, Rödel, Delfour, et al., 2017, and measured from video footage).
For the current study, the return latencies of each dolphin were averaged for each of the Near‐positive, Middle, and Near‐negative cues (instead of taking an average of all three ambiguous cues as in Clegg, Rödel, Delfour, et al., 2017).
This approach was chosen to fully investigate any associations between our one behavior variable (as opposed to multiple behaviors tested in Clegg, Rödel, Delfour, et al., 2017) and cognitive bias.
If correlations are present it would be fruitful to be able to see whether they are conserved among the different ambiguous cues, yielding as much information as possible on the link between cognitive bias and anticipatory behavior as it is clearly a species‐independent topic of interest (Wichman et al., 2012).
For each of the three ambiguous cues, the final response variable (as in Clegg, Rödel, Delfour, et al., 2017) was the percentage deviation of the ambiguous cue average return latency from the individual's average return latency from the Positive and Less‐positive (conditioned) cues.
This allowed us to control for the different swim speeds of the dolphins, and produced a measure of individual response to each ambiguous cues relative to the conditioned ones. 2.3 Behavioral data collectionDuring the three days when the final cognitive bias tests took place for each animal, behavioral data was also taken during their “free‐time” in order to assess the level of anticipatory behavior.
Focal observations of 5 min with scan sampling for behaviors every 15s (Mann, 1999) were conducted once a day within the 15‐min period before one of the five main training sessions (between 10:00 and 16:30), in order to capture an accurate reflection of anticipation for these events specifically.
These training sessions could either include the cognitive bias testing or not (see description of training sessions earlier in the methodology), but these were conducted at random times of the day and there was no way for the animal to discern the contents of the session before it started: they were only able to predict the start time using environmental and time‐delay cues.
The same person (IC) always conducted the observations using a behavioral repertoire of the two most commonly described anticipatory behaviors in the literature (Clegg, Rödel, Cellier, et al., 2017; Miller et al., 2011): surface‐looking and spy‐hopping (Table 1).
Both of these behaviors were found to be among those that increased before training sessions in two previous studies at Parc Astérix (Clegg, Rödel, Cellier, et al., 2017; Jensen et al., 2013).
For analysis, the scans of both anticipatory behaviors were summed and then converted to a percentage of total scans per observation (i.e., to correct for non‐visible scans), and are therefore described as frequencies hereafter.2.4 Data analysisStatistical analyses were done with R, version 3.1.1 (R Development Core Team, 2016).
For our first question regarding whether test criteria were maintained, we used the Fisher‐Pitman permutation test (package coin, Hothorn, Hornik, Van de Wiel, & Zeileis, 2006).
To test the associations between cognitive bias test results and anticipatory behavior frequency, we used linear regression models. p‐values were calculated by Monte Carlo sampling with 10,000 permutations, using the R package pgirmess (Giraudoux, 2016).
Such permutation tests are especially appropriate for moderate sample sizes and do not have any assumptions regarding normal distribution of residuals (Good, 2005).
The anticipatory behavior frequency represented the independent variable in three separate models with the relative return latency for the NP, M, and NLP ambiguous cues as the response variable in each.
The Bonferroni correction was not applied to p‐values because it was felt that the three variables, that is, the three ambiguous cues NP, M, and NLP, were measuring a facet of the same phenomenon and thus not truly independent (the correction should be applied when the “variables being tested are independent,” Garcia, 2004).
We also used linear models and permutation tests to investigate whether anticipatory behavior frequencies and relative return latencies from each ambiguous cue differed with the sex of the animals (included as a factor in each model).3 RESULTS3.1 Absolute return latencies for all cues
A descriptive graph showing the absolute return latencies for each dolphin over all the cues in the final testing phase (Figure 1) confirms that the animals maintained their differentiation between the conditioned cues (P and LP), with on average a difference of 2.0 s (min: 1.5 s, max: 2.4 s; Fisher–Pitman test with 10,000 permutations: Z = 11.72, p < 0.001; Clegg, Rödel, Delfour, et al., 2017).
We can also see that in general the latencies for all dolphins increase incrementally as we move from further away from the Positive and closer to the Less‐positive cue positions. 3.2 Association between anticipatory behavior and ambiguous cue return latencies
There was a positive, significant correlation between anticipatory behavior frequency and the relative return latency from the Near‐positive cue (linear regression with 10,000 permutations: R2 = 0.593, p = 0.034; Figure 2a), and a similar, although non‐significant, tendency with respect to return latency from the Middle cue (R2 = 0.410, p = 0.087; Figure 2b).
Those animals performing more anticipatory behavior also returned slower from the ambiguous cues that is, made more pessimistic decisions.
However, there was no significant correlation between anticipatory behavior frequency and return latency from the Near‐negative cue (R2 = 0.023, p = 0.749). 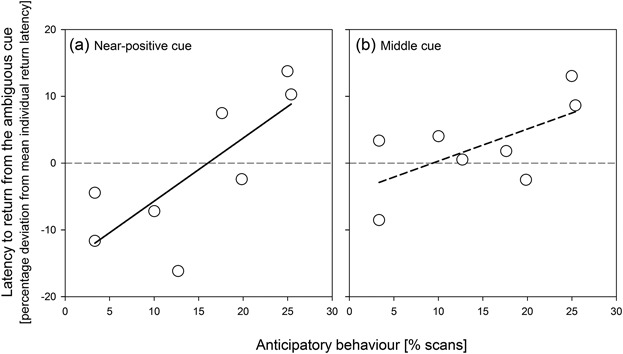 Figure 2Cognitive bias test results predicted by anticipatory behavior. (a) The dolphins' relative return latencies to the Near‐positive cue were predicted by the frequency of anticipatory behavior toward the training sessions during the third day test period: those slowest to return (pessimistic‐like judgement) also showed significantly highest frequencies of anticipatory behavior, and this same relationship was present (b) with respect to the Middle cue, although the association was a statistical tendency only. Figure 2Cognitive bias test results predicted by anticipatory behavior. (a) The dolphins' relative return latencies to the Near‐positive cue were predicted by the frequency of anticipatory behavior toward the training sessions during the third day test period: those slowest to return (pessimistic‐like judgement) also showed significantly highest frequencies of anticipatory behavior, and this same relationship was present (b) with respect to the Middle cue, although the association was a statistical tendency only.
Note that high and positive y‐axis values correspond to an pessimistic‐like judgement, and the lower and negative to optimistic‐like judgements.
See text for details on statistics. Anticipatory behavior in relation to upcoming training sessions was the combined frequencies of two behaviors, surface‐looking, and spy‐hopping (see Table 2 for definitions)
There were no significant differences between males and females with respect to the frequency of anticipatory behavior, nor their return latencies from the Near‐positive, Middle, or Near‐less‐positive cues (all p > 0.05). 4 DISCUSSIONWe found that the frequency of anticipatory behavior toward upcoming training sessions predicted cognitive bias in bottlenose dolphins.
Dolphins showing the higher frequencies of anticipatory behavior also made more pessimistic judgements in relation to the Near‐positive ambiguous cue, and tended toward the same association for the Middle cue.
Our protocol was the first application of cognitive bias testing to marine mammals and only used positive reinforcement (no negative reinforcement or punishment): the test methodology was validated by the fact that on average the dolphins continued to significantly discriminate between the conditioned cues for the duration of the test period.
Our findings are the first to provide evidence for a link between anticipatory behavior and cognitive bias in any species, both of which have been proposed as measures of animals' affective states (Mendl et al., 2009; van der Harst & Spruijt, 2007).
A recent study with laying hens predicted that as both cognitive bias and anticipatory behavior are thought to be linked to welfare, the results of concurrent testing would be significantly correlated: however, they were not able to prove this, and suggested this could be due to their methods of testing anticipatory behavior and influences of the social group (Wichman et al., 2012).
In our study with dolphins, increased anticipatory behavior was associated with slower return times, or more pessimistic judgements: significantly for the Near‐positive ambiguous cue, a tendency for the Middle cue, but not for the Near‐less‐positive position.
It is first important to address the difference in results between ambiguous cues: this is similar to findings in previous cognitive bias animal studies, where focus is placed on a significant result found with any ambiguous cue. However, in human studies and recently in the chick model, it has been shown that variation in responses to the near‐positive or near‐negative cues can, for example, differentiate between states of anxiety and depression (Mendl et al., 2009; Salmeto et al., 2011).
Therefore although we are currently unable to interpret these differences in results, the development of perhaps more meticulous testing protocols could result in very useful outcomes for animal welfare management.
Our results supported our hypothesis: we found dolphins that anticipated the training sessions more intensely also judged more pessimistically in a decision making task.
Pessimistic judgements have been associated with negative affective states in many other species and settings (Baciadonna & McElligott, 2015; Mendl et al., 2009; Roelofs et al., 2016).
According to the reward‐sensitivity theory, dolphins experiencing more negative affective states would show higher anticipatory behavior before positive events because they place a higher value on the reward to come, that is, they desire it more, perhaps due to a lack of other positive events in their environment (Spruijt et al., 2001).
Conversely, the dolphins that made more optimistic judgements may have been in more positive affective states, and thus placed less value on the upcoming reward and anticipated it less.
However, as discussed in the previous paper on this dataset (Clegg, Rödel, Delfour, et al., 2017), the design of the study means that causal relationships between the behaviors and affective states represented by the cognitive bias results can not be confirmed. Although steps were taken to reduce interference from other dolphins on the test dolphin, the fact that we tested the animals while they had free access to the rest of their group means we cannot rule out that social learning may have occurred for some animals regarding the response to cues.
However, as the half animals had the extreme left side of the arc as a positive cue and the other half had the right extreme, social learning for these cues would have had to occur between only selected dolphins. Another alternative hypothesis is that the dolphins judging pessimistically in fact viewed the feeding/training sessions as negative events (Frohoff & Packard, 1995; but note that the reward‐sensitivity theory does not apply to negative events, Spruijt et al., 2001).
Although anticipatory behavior of captive dolphins is still a very new research area, current evidence suggests that we might reject this alternative hypothesis: firstly, it has been shown with other species that the type of anticipatory behavior differs in response to upcoming positive versus negative events (e.g., Moe et al., 2006; Zimmerman, Buijs, Bolhuis, & Keeling, 2011).
Therefore if some dolphins were anticipating the training sessions in a negative light, we would not have likely seen such a significant association with cognitive biases as the anticipatory behaviors measured would have appeared differently; for example, we might have seen behaviors with links to fear or frustration (as with hens, Zimmerman et al., 2011) and more avoidance of the area where sessions started, whereas, we saw only approaching behaviors. Finally, there is some evidence from measures of physiological stress that training/show/guest interaction sessions are not likely to be viewed negatively by the animals: the breathing rate (St. Aubin & Dierauf, 2001) of bottlenose dolphins did not differ between the periods before and after show sessions (Jensen et al., 2013), and three stress hormones of belugas (Delphinapterus leucas) did not change from baseline levels during guest interaction sessions (Schmitt, St. Aubin, Schaefer, & Dunn, 2010).
As a result, we tentatively conclude that the dolphins in this study viewed the training sessions positively and that our results are thus in line with the reward‐sensitivity theory.
The anticipatory behaviors of captive dolphins studied here, surface looking and spy‐hopping, are direct products of the environment as the animals are using cues to prepare for an event that will occur in the near future (Jensen et al., 2013). Further investigations with controlled variation of environmental and management factors would help us to better understand anticipatory behaviors, as well as, for example, basic experiments with captive animals to discover whether increased anticipatory behavior means the animal is more motivated for the reward, i.e., do anticipation levels predict behavior during the reward's acquisition?
It could be that animals that strongly anticipate a positive reward, but do not exploit it once it arrives, are in poorer welfare than those highly anticipating the reward then actively exploiting it (the former situation could be similar to stereotypic behavior which has no apparent function, Mason & Rushen, 2008).
Our study has shed some light on the frequencies of anticipatory behavior in captive dolphins which might be linked to positive and negative affective states, and thus could start to fill in the gaps for this species (i.e., thresholds on the proposed models: van der Harst & Spruijt, 2007; Watters, 2014).
However, our work is only the first step and was conducted on a moderate sample size of animals: more work is needed on multiple groups to uncover the real significance and utility of anticipatory behavior measurement in captive bottlenose dolphins. ACKNOWLEDGMENTS
We would like to thank Birgitta Mercera and the Parc Astérix training team for their invaluable support and involvement in this study.
We are also indebted to Heiko G. Rödel who aided greatly in statistical analyses and development of the manuscript.
Thank you also to the two anonymous reviewers who contributed greatly to the quality of the manuscript.
This work forms part of IC's PhD project and was partly supported by CIFRE fund no. 2014/0289 and UFAW grant no. 22‐14/15.CONFLICTS OF INTERESTIsabella Clegg and Fabienne Delfour declare that they have no conflict of interest.ETHICAL APPROVALAll applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
The experimental protocol followed the ASAB/ABS Guidelines for the Use of Animals in Research (Association for the Study of Animal Behaviour, 2012), and was accepted by the scientific and animal welfare committees of Parc Astérix. ------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------- Tot zover het naar ik aanneem meest wetenschappelijke artikel dat ooit op Circuswereld is vreschenen! Gelukkig staan er afbeeldingen bij, dat maakt het wat minder saai en leesbaarder...... Eén tabel ontbreekt, want bleek niet kopieerbaar. En niet alle tussenkoppen zijn in dezelfde grootte, dat komt misschien nog wel...... Aardig zijn wel de verwijzingen naar (naar ik aanneem) Nederlandse wetenschappers die eerdere onderzoekingen deden.
|
|